Формально образование белковой макромолекулы можно представить
как реакцию поликонденсации α-аминокислот:
При взаимодействии двух молекул α-аминокислот происходит реакция
между аминогруппой одной молекулы и карбоксильной группы - другой. Это приводит
к образованию дипептида (часть IV, раздел 4.3),
например:
Следует отметить, что в искусственных условиях
(вне организма) две различных аминокислоты могут образовать 4 изомерных дипептида
(в данном случае - глицилаланин, аланилглицин, аланилаланин и глицилглицин).
Из трех молекул α-аминокислот (глицин+аланин+глицин)
можно получить трипептид:
Аналогично происходит образование тетра-, пента- и полипептидов.
Количество изомерных пептидов резко
возрастает с увеличением числа участвующих в их образовании неодинаковых
α-аминокислот. В живом организме (in vivo)
биосинтез полипептидов (белков) с заданной природой последовательностью α-аминокислотных
остатков направляют дезоксирибонуклеиновые кислоты (ДНК).
Непосредственно его осуществляют рибонуклеиновые кислоты (РНК информационные,
транспортные, рибосомные) и ферменты.
Для искусственного получения олиго- и полипептидов заданного строения
применяются специальные приёмы пептидного синтеза: блокирование
(защита) одних функциональных групп и активация других.
При синтезе полипептидов, содержащих большое число аминокислотных остатков,
требуется проведение множества реакций, которые сопровождаются операциями по
выделению и очистке продукта на каждой стадии. В классическом синтезе это
сопряжено не только с большими затратами времени, но и с потерями вещества,
поэтому при завершении эксперимента удается получить лишь ничтожно малые количества
конечного продукта.
Для решения этих проблем был предложен твердофазный синтез пептидов
на полимерной матрице (Мэррифилд, 1962). В настоящее время созданы автоматизированные
приборы (синтезаторы), которые в запрограммированной последовательности осуществляют
все необходимые операции. Если в классическом синтезе для присоединения одной
аминокислоты требуются дни и даже недели, то условия твердофазного синтеза
позволяют присоединить 6 аминокислот в сутки.
Молекулярные массы различных белков (полипептидов) составляют от 10 000 до нескольких миллионов. Макромолекулы белков имеют стеререгулярное строение, исключительно важное для проявления ими определенных биологических свойств.
Несмотря на многочисленность белков, в их состав входят остатки не более 22 α-аминокислот.
Разнообразные функции белков определяются α-аминокислотным составом и строением их высокоорганизованных макромолекул.
Выделяют 4 уровня структурной организации белков.
Первичная структура
- определенная последовательность α-аминокислотных остатков в полипептидной цепи.
Вторичная структура
- конформация полипептидной цепи,
закрепленная множеством водородных связей между группами N-H и С=О. Одна из моделей
вторичной структуры - α-спираль.
Другая модель - β-форма ("складчатый лист"), в которой преобладают межцепные (межмолекулярные) Н-связи.
Третичная структура
- форма закрученной спирали в пространстве, образованная главным образом за счет дисульфидных мостиков -S-S-, водородных связей, гидрофобных и ионных взаимодействий.
Четвертичная структура
- агрегаты нескольких белковых макромолекул (белковые комплексы), образованные за счет взаимодействия разных полипептидных цепей

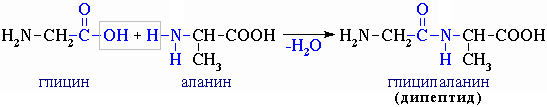





){kind=link}